
First, we highlight the recent research(Leipe, 2017) that focus on the role of the Okhotsk culture in dispersing a different type of barley from that cultivated by the Yayoi people south of Hokkaido;
Second, we show that this dispersal pattern by the Okhotsk People in Hokkaido is corroborated by the hypothesized global patterns and paths of the dispersal of barley strains in East Asia/Eurasia.,
Third, the revealed barley dispersal paths(Lister et al., 2018) especially the divergence of the northern and southern routes support our recent understanding of population substructure and heterogeneity of the genetic components of Jomon and Ainu ancestry which has led to a call to review the dual structure model, in the light of the evidence that supports northern migrations into Hokkaido that have formed part of the ancestry of the modern Ainu and Hokkaido Jomon population.
These dispersals were not only cultural but demic as well, and the genetic components exist still in the Northeast of Japan. It might be time to ditch the dual structure model in favor of a multiple migration model (or at least a more nuanced model) for the peopling of the Japanese archipelago, combining more comprehensive evidence from interdisciplinary fields.
…
Below, we run through excerpts of the relevant literature.
I. Christian Leipe et al., Barley (Hordeum vulgare) in the Okhotsk culture (5th–10th century AD) of northern Japan and the role of cultivated plants in hunter–gatherer economies, 2017 excerpts follow below:
This paper (archaeobotanical/paleobotsnical approach) by Leipe throws light upon the changing subsistence strategies of the Hokkaido Jomon, the Satsumon, as impacted upon by the arrival of the Okhotsk people. By focusing on the dispersal patterns of barley into Hokkaido, the study also helps us understand the migration patterns of the Okhotsk people from the north into Hokkaido. Two types of barley, alongside of broomcorn and foxtail millet, were grown in Hokkaido as crops complementary to the foods obtained through foraging by the Okhotsk people who remained complex hunter-fisher-gatherers unlike the agricultural Yayoi people to the south.
Relevant arguments and supporting evidence connecting the now-extinct Okhotsk culture with barley cultivation are excerpted below:
This paper discusses archaeobotanical remains of naked barley recovered from the Okhotsk cultural layers of the Hamanaka 2 archaeological site on Rebun Island, northern Japan. Calibrated ages (68% confidence interval) of the directly dated barley remains suggest that the crop was used at the site ca. 440–890 cal yr AD. Together with the finds from the Oumu site (north-eastern Hokkaido Island), the recovered seed assemblage marks the oldest well-documented evidence for the use of barley in the Hokkaido Region. The archaeobotanical data together with the results of a detailed pollen analysis of contemporaneous sediment layers from the bottom of nearby Lake Kushu point to low-level food production, including cultivation of barley and possible management of wild plants that complemented a wide range of foods derived from hunting, fishing, and gathering.
To our knowledge, the archaeobotanical assemblages from the Hokkaido Okhotsk culture sites highlight the north-eastern limit of prehistoric barley dispersal. Seed morphological characteristics identify two different barley phenotypes in the Hokkaido Region. One compact type (naked barley) associated with the Okhotsk culture and a less compact type (hulled barley) associated with Early–Middle Satsumon culture sites. This supports earlier suggestions that the “Satsumon type” barley was likely propagated by the expansion of the Yayoi culture via south-western Japan, while the “Okhotsk type” spread from the continental Russian Far East region, across the Sea of Japan. After the two phenotypes were independently introduced to Hokkaido, the boundary between both barley domains possibly existed ca. 600–1000 cal yr AD across the island region.
Researchers are reconstructing the dispersal of barley and other crops of the Southwest Asian agricultural package out of their ‘core area’. In addition to rapidly dominating the economy of peoples in Neolithic Europe, barley spread eastward across the Asian mainland. Barley arrived in the greater Indus valley region and southern Turkmenistan by ca. 6000 cal yr BC [10], to monsoon-dominated India after 3000 cal yr BC [10] and the mountain foothills of Kazakhstan in Central Asia by 2200 cal yr BC [11], to Central China [12], the Tibetan Plateau [13] and the Korean Peninsula [14] around 1500 cal yr BC, and to the Primor’e Region in the Russian Far East (RFE) after 400 cal yr BC [15].
The only archaeobotanical data we have for its spread into the northwest Pacific islands comes from the Japanese archipelago, with no studies thus far conducted in Sakhalin and the Kurils [16]. Japanese scholars claim that preserved barley grains have been sporadically recovered from archaeological excavations on Honshu and Kyushu islands (see [17] and references therein) from contexts dating to the Middle–Final Jomon period (ca. 3500–100 cal yr BC, according to [18]). However, intensive and widespread cultivation of barley, as well as rice and wheat does not appear to have been present prior to the Initial Yayoi period on Kyushu (ca. 1000–900 cal yr BC, see [18] and references therein) and around 100 cal yr BC (according to [18]) on north-eastern Honshu. Thus, the Jomon/Yayoi transition in central and southern Japan marks the switch from an economy dominated by foraging to one based on agricultural production. In contrast, the Hokkaido Region (Fig 1C) was not affected by these changes in subsistence. People in northern Japan, similar to those in Greenland, Arctic regions of Asia, and the American West Coast, remained “complex” hunter–fisher–gatherer well into the historic period [19]. Local Jomon populations of Hokkaido continued a foraging lifestyle [20] until the middle of the 1st millennium AD when they were replaced by Okhotsk cultural communities in the north and by Satsumon cultural communities in the central and the southern parts of the island [21]. Both of the latter cultures are commonly identified as hunter–fisher–gatherers [19]; however, their archaeological remains show evidence for the use of metals and the cultivation of crops [8]. The extent of their productive economy has not been fully studied. To date, the only clear evidence for the cultivation of barely and other crops by Satsumon people in the late 1st millennium AD comes from a single excavation site, which is located in the municipality of Sapporo [8]. There are more data showing that their contemporary neighbours to the north, the Okhotsk culture in Hokkaido, were cultivating both broomcorn (Panicum miliaceum) and foxtail (Setaria italica) millet and barley….
These well-preserved barley specimens from Rebun Island are accurately dated and morphometrically described. We also compiled a new morphological dataset of archaeological barley grains including the Hamanaka 2 record as well as published and unpublished records from other sites in the wider study region. The morphological dataset is further used to discuss dispersal pathways to extend the existing “map” of chronological and geographical spread of domesticated barley throughout East Asia.
The Okhotsk Culture
The people of the Okhotsk archaeological culture are regarded as a hunter–gatherer society with an economy that strongly relied on marine resources. They occupied a widespread maritime environment, mainly along the southern and eastern littoral margins of the Sea of Okhotsk including northern and north-eastern Hokkaido (see Fig 1C for archaeological site distribution), Sakhalin Island, and the Kurils (Fig 1A). In the Hokkaido Region, the peak of the Okhotsk cultural occupation dates from the 6th to the 8th century AD (see [26] and references therein). Based on pottery style, the “Okhotsk cultural sequence” in northern Hokkaido is divided into three chronological stages comprising the (1) Susuya culture (2nd–5th century AD), which is often referred to as incipient or Proto-Okhotsk, (2) the Towada, Kokumon, Chinsenmon, Haritsukemon, and Somenmon cultures (6th–8th century AD) regarded as the main stages, and (3) the Motochi culture (9th–10th century AD) as the final stage [27]. While Okhotsk cultural traits persisted through the Tobinitai period in eastern Hokkaido until the 12th century AD, replacement or assimilation of the Okhotsk culture in northern Hokkaido by Satsumon/Proto-Ainu populations originating from the central and southern areas of Hokkaido was completed by the end of the 10th century AD [26].
Archaeologists believe that the Okhotsk culture people migrated to Hokkaido from the north (i.e. Sakhalin Island), first occupying Rebun and Rishiri islands as well as the northern tip of Hokkaido and subsequently dispersing eastwards along the coast of the Sea of Okhotsk [28]. Results of archaeological and genetic studies suggest that the Okhotsk population probably originated from the lower Amur River basin (e.g. [29–31]). The population spread onto the islands bordering the Sea of Okhotsk, which is believed to have been due to socio-political conflict [31]. There is also evidence for the onset of cooler climatic conditions in the lower Amur River basin around the end of the 1st millennium BC [32, 33]. These climate changes may have played a role in the southward spread (ca. 500 AD) of these people to Hokkaido [34] and their later absorption/replacement (by ca. 1000–1200 cal yr AD; [26, 27]).
A defining trait of the Okhotsk culture is its subsistence strategy, traditionally thought to be a specialised system of marine resource extraction [26, 35]. This is reflected by the geographic distribution of sites along coastal regions (Fig 1C) and confirmed by archaeological studies of faunal remains and tool assemblages, which indicate intensive marine hunting, fishing, and gathering activities (e.g. [31, 36, 37]). Nitrogen stable isotope studies on human remains also point to a diet with high proportions of protein derived from marine organisms (e.g. [35, 38, 39]). Analysis of human bone collagen revealed a relative contribution of marine protein in the range of 60–94% for individuals from Rebun Island [38] and 80–90% for individuals from eastern Hokkaido [39]. However, there is enough evidence to suggest that the diet of the Okhotsk people may have been much more diverse than the isotopic data imply. People likely supplemented the maritime resources with terrestrial mammals such as deer, fox, rabbit, and marten [37]. Cut marks on bones from domesticated dogs suggest that they were also part of the diet [36], and remains of domestic pigs are limited to northern Hokkaido [26]. In addition, there is evidence for the use of edible wild plants including Aralia (spikenard), Polygonum (knotweeds), Actinidia (Chinese gooseberry), Vitis (grapevines), Sambucus (elderberry), Empetrum nigrum(crowberry), Rubus sp. (blackberry), Phellodendron amurense (Amur corktree), and Juglans (walnut). Furthermore, as already noted, broomcorn millet, foxtail millet, and barley grains have been recovered from sites in this cultural horizon (see [26] and references therein). Admittedly, we know very little about the role of any of these plants in the economy, or whether the crops had a dietary or ritual function [22]. …
Our pollen-derived reconstruction of the local vegetation suggests stronger human impact on the island vegetation between ca. 400 and 900 cal yr AD, corroborating the archaeological data for the regional Okhotsk culture occupation phase.
The results of the pollen analysis also indicate that the reconstructed phase of intensified human impact was superimposed onto a long-term climatic cooling trend, suggested by decreasing percentages of Quercus and Ulmus. Spanning the entire record, both of these thermophyllous taxa show a continuous gradual decline from roughly 10% and 5% at the bottom of the record to about 5% and 1.5% at the top, respectively. This trend likely indicates a progressive cooling in the study region in line with the late Holocene trends of Northern Hemisphere summer insolation [57] and published palaeoclimatic records from the region (e.g. [58, 59]). …
There is evidence that human migrations from the north have played an important role in the prehistory of Hokkaido and other parts of the Japanese archipelago. This includes the intrusion of Siberian Palaeolithic hunter–gatherer groups around the Late Glacial Maximum (ca. 20,000 cal yr BP; [60]) and immigration ca. 15,000 cal yr BP, with the latter introducing microblade technologies on Hokkaido and Honshu [61, 62]. While they have not been taken into account for a while (e.g. [63]), recent anthropological studies (e.g. [64]) stress the role of migration from northern regions via Hokkaido also in view of the origins of the Neolithic Jomon culture. The most recent southward movement of prehistoric populations into the northern and north-eastern coastal areas of Hokkaido was that of the Okhotsk culture around the middle of the 1st millennium AD [26]. Though, the Okhotsk groups inhabited a large area along the southern and eastern margins of the Sea of Okhotsk, most of our current knowledge has been derived from archaeological materials recovered in the Hokkaido Region. The archaeobotanical record from the Hamanaka 2 site presented in this study allows for greater insight into the use of plants by the Okhotsk people on Rebun Island (Fig 1D). Calibrated ages (95% confidence interval) of directly dated barley remains from five archaeological layers (IIIa–e) suggest that the crop was used at the site between 430–960 cal yr AD (Table 2) or at a 68% confidence interval between ca. 440 and 890 cal yr AD. This time period roughly corresponds to the late Susuya through mid-Motochi stages spanning between the 5th and 10th century AD [27], thus covering the Okhotsk culture settlement phase in northern Hokkaido as indicated by previous archaeological studies. Given the age of the oldest barley seed F2014-037-003 (440–600 cal yr AD, 68% confidence interval; Table 2), the Hamanaka 2 layer IIIe, together with the single dated grain (428–573 cal yr AD, 95% confidence interval; [65]; S3 Table) from the Oumu site (no. 29 in Fig 1C) represents, the earliest well-documented record of domesticated barley in the Hokkaido Region. The only carbonised barley grain recovered in Hokkaido was collected from the Epi-Jomon level of the K135–4 Chome site within the city of Sapporo [66]. This single barley seed has not been directly dated and its proposed age of ca. 200–400 cal yr AD should be viewed with caution.
The presence of barely at the Hamanaka 2 site appears contemporaneous with a phase of enhanced human-induced vegetation disturbance on Rebun Island as indicated by the pollen record from Lake Kushu (Fig 5). During this time (with a maximum ca. 550–800 cal yr AD), the pollen record shows a decrease in the abundance of arboreal pollen, suggesting deforestation and greater openness of the landscape compared to the preceding and subsequent periods, which are more or less coeval with the Epi-Jomon and Proto-Okhotsk (Susuya) cultures (ca. 100 cal yr BC–500 cal yr AD) and Proto-Ainu culture (ca. 950–1600 cal yr AD), respectively ([26, 27]; Fig 5). The results of the local vegetation reconstruction clearly indicate enhanced human activities on Rebun Island during the main phase of the Okhotsk presence there. On the other hand, reduced impact is evidenced during the Epi-Jomon phase and the time of cultural shifts towards the Classic Ainu period, which may be explained by reduced population size and/or a different pattern of resource exploitation. Regarding the Epi-Jomon, this would conform to identified traits like short-term occupations, high mobility, and low complexity [66, 67]. Ohyi [28] suggests that by the time of the disappearance of the Okhotsk culture at the end of the Motochi stage, the Satsumon people spread into northern Hokkaido, including Rebun Island and neighbouring Rishiri Island. It appears, at least for Rebun Island, that these Satsumon groups weakly impacted the island’s vegetation, which was leading to the recovery of local fir forests. Fewer human activities at Hamanaka 2 are also suggested by the absence or quantities of archaeological materials (unpublished data) associated with the Satsumon culture in the subdivisions of layer II.
Rebun Island is well-known for its Okhotsk culture sites, representative for the northern Hokkaido domain. Here, the presence of the Okhotsk groups continued into the Motochi stage (9th–10th century AD) at a time when the Okhotsk sites in northern Hokkaido became abandoned [28]. However, on Rebun Island the presently discussed Hamanaka 2 assemblage contains the only barley thus far recovered (T. Amano, personal communication). This might be due to a lack of systematic sampling and water flotation at other sites on the island. We identified the recorded barley as naked barley, which is far more commonly found than hulled barley in East Asia [68]. Barely was consumed at the site over a ca. 500-year period throughout the main stages of the Okhotsk culture (Fig 5). Barley was significant and had a long-term role in diet during the peak of the Okhotsk culture in the region. The use of barley is also evident at other sites in north-eastern Hokkaido (Fig 1C), being assigned to the late phase (8th–9th century AD) of the Okhotsk culture [69]. In addition, remains of foxtail and broomcorn millet are reported from several excavations [69]. Japanese palaeobotanists have argued that these crops were used for ritual purposes (e.g. [22]); however, this is hard to defend seeing that they appear in so many domestic contexts across such a large time period. The grains likely supplemented a mixed economic system that relied heavily on wild coastal resources. Although, an alternative hypothesis is that these crops were used to produce alcohol [70]. Another important question to ask is whether barley was obtained by trade or cultivated locally by the Okhotsk people or a combination of both. At Hamanaka 2, no tools related to barley cultivation or processing have been found and no tests of organic residue on pottery have been conducted yet to support either use. Yamaura [71] argues that the axe- and hoe-shaped bone tools unearthed in eastern and northern Hokkaido Okhotsk sites (including Kafukai A, southern Rebun Island) represent evidence for crop cultivation. However, this is not conclusive evidence, since such tools might have had other purposes like collecting wild plants and digging dwellings and/or storage pits.
One reason why some scholars have been hesitant to accept that barley and other cereals were dietary supplements may be that the Okhotsk are generally regarded as a specialised hunter–gatherer culture with a subsistence strongly focusing on maritime food resources. This traditional view of a coastal foraging society has been bolstered by recent human bone isotope studies (e.g. [35, 38, 39]), which revealed a high proportion of absorbed protein derived from marine resources of up to 94% and 90% in northern and eastern Hokkaido, respectively [39]. However, our findings together with results of previous studies illustrate that the Okhotsk relied on a wide range of natural and domesticated foods. Besides the suggested strong focus on marine collecting, fishing, and mammal hunting, the Okhotsk people appear to have employed a broad spectrum of wild terrestrial plant fruits and root tubers (this study, Table 3; [26] and references therein), hunted a variety of terrestrial mammals [37], and also maintained domesticated dogs [36] and pigs [26] as part of their food economy. Indications for plant maintenance also comes from the increase in Lysichiton type pollen in the Lake Kushu pollen record (Fig 5) and carbonised Cyperus sp. root tubers in the Hamanaka 2 flotation samples (Table 3). Both taxa represent plants growing in swampy environments around Lake Kushu, which include edible parts and provide nutritious food. It is conceivable that the local Okhotsk people exploited these plants and even maintained their growth and productivity using suitable tools for tilling as found in contemporaneous cultural strata on Rebun Island [71].
Given the combination of foraging, animal husbandry and the use of barley and other cereals over a wide spatio-temporal array, crop cultivation as a supplementary portion of Okhotsk subsistence seems more plausible. This case study further augments existing examples of (“complex”) hunter–gatherers, occupying the “middle ground” which separates hunting–fishing–foraging societies exclusively depending on wild food resources and agriculturalists with a major focus on managing and producing domesticated plants and animals (e.g. [72, 73]). It has been noted that this middle ground territory is highly complex. As Smith [9] puts it, “this territory between hunting–gathering and agriculture is turning out to be surprisingly large and quite diverse; it has also proven to be quite difficult to consistently describe in even the simplest conceptual or developmental terms”. Smith [74] built his concept of ‘low-level food production’ on earlier observations by Braidwood and Howe [75] as well as Flannery [76], all of whom use the term “incipient cultivation” to describe intermediary strategies between foraging and farming. Many other ethnographers and archaeologists have subsequently noted that there is a wide range of diversity in human economic systems; notably, Boserup [77] points out that the range of land-use strategies reflect an equally broad range of human adaptive economies. There have been different approaches to define the middle ground landscape. Following the conceptual framework of Smith [9], who identified low-level food production (<50% annual caloric budget from domesticates) relating to tended wild plants and/or cultivated/managed plants. The Okhotsk culture may be confidently placed somewhere between the traditionally accepted boundaries between foraging and agriculture. The same view is taken by Crawford [8] who explicitly assumes that the Okhotsk people themselves cultivated barley and millet. Given the evidence for dog and pig husbandry and the cultivation or exchange of barley, the Okhotsk epitomises the complexity and diversity of the middle ground economy. Smith [9] illustrates that for many prehistoric societies this in-between space did not mark a brief transitional phase towards agriculture, but a state of stable long-term (often over several millennia) or even permanent economies combining “low-level production” of domesticates and a major focus on wild resources. Such more or less stable “mixed” subsistence economies are also evidenced in a number of modern and historic hunter–gatherer groups and are mostly characterised by either the adoption of cultivation or the exchange of domesticates (e.g. [78] and references therein). In light of the present evidence, it appears that also the Okhotsk people employed such socio-political and economic strategies.
Thus far, the ongoing debate about the middle ground landscape has either been centred on early “mixed” subsistence economies (e.g. in New Guinea [7] or eastern North America [4]) or on recent ethnographically observed cultures (e.g. the Mikea people of Madagascar [79] or the Agta of the Philippines [80]). In this regard, the adoption of domesticates by the Okhotsk people, which occurred in relatively recent prehistory, adds particular value as it bridges the gap between the two foci. Another specific feature of the Okhotsk subsistence strategy is the process of adopting already fully domesticated plants that were, by this time, widely used as staple crops in agrarian societies across Eurasia. In the Okhotsk culture, however, the incorporation of barley and millets does not appear to have had significant socio-economic effects. While the subsistence economy continued to be based on foraging, the society remained egalitarian and “group-oriented” [26]. Similar observations are reported from other regions like Japan as well as Island Southeast Asia and Melanesia. The spread of domesticated rice through the latter two regions appears to have started at the beginning of the 1st millennium BC [81]. However, in most regions, rice remained a minor supplementary crop in subsistence systems mainly based on vegeculture (e.g. taro, banana, and sago production) and foraging [82] until the middle of the 1st millennium AD [83]. In Japan, the oldest botanical remains of domesticated broomcorn and foxtail millet, barley, and rice date between the Middle and Late Jomon periods [17], thus may also indicate an early (pre-Yayoi) introduction of domesticated cereal crops from outside the archipelago as minor subsistence supplements not signifying a fundamental change in dietary pattern. All these examples immediately prompt the urgent question of what social or environmental conditions govern the decision of vegeculturalists and/or foragers to adopt domesticated seed crops? While progress has been made in addressing this central issue in anthropological and archaeological research (e.g. [84]), a more universal conceptual framework, if at all possible, has thus far not been suggested.
Another feature of the analysed flotation samples that raises questions is the abundance of Rhus/Toxicodendron seeds. There is ongoing debate about the taxonomic standing within these genera; while some scholars have treated Toxicodendron as a separate genus, others have treated it as a subgenus of Rhus (see [85] and references therein). In some regions plants of the Rhus complex have a long history as a medicine or spice [86]. Significant numbers of Rhus/Toxicodendron seeds were recovered from Jomon sites in southern Hokkaido [8]. In this record at least two Rhus/Toxicodendron species were found, one of which was identified as Toxicodendron vernicifluum (lacquer tree), which is related to lacquer production [8] and medical effects [86]. Lacquer items were major trade goods and symbols of authority throughout the past four millennia in East Asia. An example showing the value of such items from neighbouring regions are lacquer artefacts recovered from elite tombs at major urban centres such as Erlitou [87] in China dating back to the period that most Chinese scholars call the Shang (late second millennium BC). Today, three species of Rhus/Toxicodendron grow in the Hokkaido Region including Toxicodendron trichocarpum (syn. Rhus trichocarpa), Toxicodendron orientale (syn. Rhus ambigua), and Rhus javanica [88]. The Toxicodendron species (i.e. T. vernicifluum, T. trichocarpum, and T. orientale) contain toxic substances that can cause severe allergic dermatitis by direct contact with its plant parts or exposure to smoke or fumes from burning plant parts. Based on the pericarp cross section structure, Yoshikawa and Ito [89] have proposed a method to distinguish carbonised seeds of T. vernicifluum, T. trichocarpum, T. orientale, and R. javanica. However, application of this method failed to differentiate the Rhus/Toxicodendron seeds extracted from the Hamanaka 2 site flotation samples. Further research is needed to facilitate robust identification of carbonised Rhus/Toxicodendron seeds to species level and to understand the meaning (e.g. medical effects such as anti-inflammatory, antimicrobial, and antiviral [86]) of Rhus/Toxicodendron species for prehistoric people.
The adoption and dispersal of domesticated plants is a central topic of archaeological research and an important issue in understanding agricultural developments in different parts of the globe. So far, the assemblages from Okhotsk and Satsumon sites in Hokkaido represent the north-eastern edge of prehistoric barley dispersal across Asia. The upper end (600 cal yr AD) of the calibrated age range (68% confidence interval) of the oldest barley seed contained in the dated Hamanaka 2 sample set is coeval with the onset of the Satsumon culture (beginning 7th century AD), which is believed to have arisen from the Tohoku Region (Fig 1A) Yayoi culture populations driven to Hokkaido by expansion of the first Japanese state [8]. Therefore, the most straightforward inference would be that the barley used by the Okhotsk was derived from Satsumon groups spreading into the central and south-western part of Hokkaido. In fact, previous palaeobotanical work points to a different origin that is further emphasised by the Hamanaka 2 barley seed inventory. In previous studies, Japanese scholars claimed to have identified a short and a long barley type at Okhotsk and Satsumon culture sites in the Hokkaido Region, which they assigned to the crop’s naked and hulled form, respectively (see [69] and references therein). Based on this differentiation and seed morphology, Yamada and colleagues (e.g. [22, 23, 69, 90]) have hypothesised that Okhotsk barley originated from neighbouring regions on the Asian mainland. They found that the highly compact (naked barley) specimens extracted from four Okhotsk culture sites (no. 17–20 in Fig 1C, S3 Table) are distinct from the slimmer (hulled) barley (dated to 8–10th century AD) used by Early and Middle Satsumon groups, but similar to grains identified as naked barley found in the early Iron Age to medieval (ca. mid-1st millennium BC–early 13th century AD) sites in southern Primor’e (RFE). Their morphological comparison of barley grains is based on a length/width ratio dataset. Here, we review their approach by supplementing the available datasets, with eleven (partly unpublished) assemblages from the RFE, three records from northern Tohoku, and the measurements of the Hamanaka 2 site barley (S3 Table). Although naked barley is in general more compact than its hulled counterpart, quantitative morphological comparison allows for objective qualification of the recorded archaeological barley and may provide further confirmation for the proposed Okhotsk barley origin. The Okhotsk barley, which appears to be the most compact of all gathered records (Fig 4, S3 Table), in terms of shape is more similar to its counterparts found in both earlier and later (ca. 850 cal yr BC–1000 cal yr AD) sites located along coastal regions across the Sea of Japan (i.e. southern Primor’e, Fig 1C) and in the Amur River basin (Fig 1A), as opposed to grains recovered from contemporary Satsumon culture sites situated in central and southern Hokkaido. Both the Satsumon and Heian grains, which are morphologically similar to each other, appear generally longer and narrower than those used by the Okhotsk culture. This corroborates Crawford’s hypothesis that the Satsumon culture emerged from the Tohoku Yayoi culture [8], which, when being forced to migrate to Hokkaido, brought their barley with them. Alternatively, this similarity may at least suggest cultural interactions between Satsumon populations and communities in Tohoku. The results also corroborate the hypothesis that the Satsumon type barley represents hulled barely and that the naked Okhotsk barley originated in the continental RFE Region. The minor discrepancies in L/W ratios between the naked barley from Okhotsk sites and sites in the RFE (Fig 4) may reflect morphological differences commonly existing among landrace varieties of crops [91] or may be the result of different environmental conditions or irrigation [92]. This means that the barley used by the Okhotsk culture was either derived by exchange with continental populations (e.g. Mohe culture, Bohai State) from across the Sea of Japan or was brought along and cultivated by the Okhotsk culture from their region of origin (i.e. the lower Amur River basin). Unfortunately, no palaeobotanical studies have been conducted in the lower Amur River basin or on Sakhalin Island, which would have allowed us to trace back potential pathways of a southward barley introduction to Hokkaido. Further evidence for the existence of two barley phenotypes in Hokkaido comes from sites, which post-date the main phase of Okhotsk culture. Both types—the compact (naked) barley found at Okhotsk culture sites and the slim (hulled) barley found at Early to Middle Satsumon sites—are represented in archaeobotanical records from this time indicating that Okhotsk type naked barley cultivation/use continued during times of acculturation (i.e. Tobinitai culture) and the Late Satsumon stage [23]. In sum, these findings suggest that naked and hulled barley spread eastward through Asia and were introduced into the Japanese archipelago via different routes. While the area where hulled barley is recovered parallels the distribution of the Yayoi culture (south-western and central Japan) and the Satsumon culture (south-western and central Hokkaido), naked barley possibly propagated from Primor’e and adjacent regions during the Okhotsk culture spread, into Sakhalin Island and northern/north-eastern Hokkaido (Fig 1A). After the two barley phenotypes were independently introduced to Hokkaido, the boundary between both barley domains (Fig 1A) possibly existed for about 400 years across the island region until the beginning of the assimilation/replacement of Okhotsk populations by the Satsumon culture groups (ca. 1000 cal yr AD; [26, 27]).
Further evidence for a close relationship between the Okhotsk culture and Iron Age/medieval populations from the East Asian continent comes from analogies in subsistence economy as described by Sergusheva and Vostretsov [15]. Like the Okhotsk, the Yankovskaia culture (ca. 850–350 cal yr BC; [15]), which inhabited the coastal regions of today’s northern Korea and southern Primor’e, based its subsistence mainly on a wide range of marine resources. Remains of millets (broomcorn and foxtail) and naked barley found at the majority of sites suggest that these domesticates also played a role in diet there. Primor’e Region, in particular, saw a subsequent two-step advance in agricultural practices by the introduction of additional crops including wheat (Triticum aestivum/compactum), hemp (Cannabis sativa), and legumes during the Krounovskaia/Tuanjie culture (ca. 500 BC–200/300 cal yr AD; [15]) and cultigens including hulled barley (Hordeum vulgare), soy beans (Glycine max), and buckwheat (Fagopyrum esculentum) along with the establishment of early states (e.g. Bohai State) after the middle of the 8th century AD. Despite this progress in agriculture, hunting, fishing, and gathering has continuously been an additional part of the diet. In addition, pig and dog breeding is evidenced during the Yankovskaia culture and the Krounovskaia/Tuanjie culture [71, 93, 94]. These subsistence traits place the above mentioned cultures of coastal RFE and northern Korea also in Smith’s middle ground [9], thus somewhere between hunting–fishing–foraging and agriculture. Systematic water flotation has been practiced for over 30 years in this region, providing archaeobotanical remains from numerous Neolithic–Middle Age sites across Primor’e. These data show that during the Yankovskaia culture, millet and barley cultivation was not the main part of the food economy, and was probably not practiced at every archaeological site [15]. Although crop cultivation seems to have been intensified by the Krounovskaia/Tuanjie culture, there is evidence that these groups, likely due to climatic cooling at the end of the 3rd century AD, partly gave up agricultural practices and re-intensified the exploitation of wild resources [95]. This, on the one hand, emphasises that the transformation towards agriculture is not necessarily a unidirectional progression, as it was once regarded, but is a reversible process. On the other hand, it suggests that crops probably had a long-term utility as complementary foods. This might apply to Okhotsk groups, which retained their once-adopted (low-level) agricultural food production as they migrated and adapted to the maritime landscapes of the Sea of Okhotsk.
Conclusions
The archaeobotanical assemblage from Okhotsk cultural layers at the Hamanaka 2 site (northern Rebun Island, Japan) contained charred grains of compact naked barley. Direct radiocarbon dating indicates long-term use of barley at the site over a period of about 500 years. Together with the finds from the Oumu site, the data that we present marks the oldest well-documented evidence for the use of barley in the Hokkaido Region. Due to the broad error ranges of the calibrated radiocarbon dates of the oldest seed remains (428/440–573/600 cal yr AD, 68% confidence interval), more precise ages cannot be defined at this time. However, it is conceivable that the people of the Okhotsk culture were using this crop since they first arrived in the Hokkaido Region (ca. 500 cal yr AD). Accordingly, barley introduction by the Okhotsk culture would pre-date its adoption or introduction by Satsumon populations by at least a century, which may speak against the hypothesis that barley was introduced to northern Hokkaido by the more agrarian south.
The macrobotanical remains of barley are not enough evidence to argue for cultivation at the site, as opposed to the importing of grains from elsewhere. However, axe- and hoe-shaped bone tools found at nearby sites were likely farming implements and do support the possibility of local cultivation. In addition to low-level cultivation, the archaeobotanical data also suggests that wild plant management was conducted by the people of Rebun Island. The pollen record from Lake Kushu indicates significant local vegetation disturbance (i.e. deforestation) concurrent with the barley record at nearby Hamanaka 2. This reconstructed landscape patchiness may point to land clearance for small-scale crop cultivation. In view of the major role of marine food resources indicated by bone isotope studies, it seems likely that cultivated crops were used as supplementary food or for brewing beer. While future studies will clarify the role of barley in the economy in this region, it does seem clear that there was some kind of low-level crop production, which was complemented by wild foods. This rather recent example of incorporating domesticated crops into a foraging society adds to the ongoing debate about defining the middle ground landscape and further emphasises its great complexity. Moreover, it highlights the need for future studies to better understand what conditions control the adoption of agricultural practices by persistent non-agrarian societies, especially if such adoption seems to be economically inappropriate. A broad array of underlying factors should be considered, which may comprise climate, population pressure, social status, ritual practices, trade activities and so on.
So far, the archaeobotanical assemblages from the Hokkaido Okhotsk culture sites highlight the north-eastern limit of prehistoric barley dispersal. Seed morphological characteristics identified two different barley phenotypes, which were likely independently introduced to the Hokkaido Region. One highly compact type (naked barley) associated with the Okhotsk culture and a less compact type (likely hulled barley) that is evident in Early–Middle Satsumon culture sites. The much more comprehensive dataset presented in this paper supports earlier suggestions that the “Satsumon type” barley was likely propagated by the expansion of the Yayoi culture from south-western Japan towards north-eastern Japan, while the “Okhotsk type” spread from the continental RFE Region, across the Sea of Japan. Although Okhotsk populations may have obtained barley by exchange, there is growing data that suggest that they cultivated naked barley locally, which they introduced directly from their region of origin (i.e. the lower Amur River basin) via Sakhalin. To further verify this hypothesis, additional palaeobotanical studies on materials from archaeological sites in these areas are essential. Nevertheless, based on existing palaeobotanical evidence, we conclude that the Okhotsk culture represents one element of the long-term and spatially broader Holocene hunter–gatherer cultural complex (including also Jomon, Epi-Jomon, Satsumon, and Ainu cultures) of the Japanese archipelago, which may be placed into Smith’s [9] middle ground subsistence strategy. This middle ground domain may chronologically include the groups dating to the Neolithic–Iron Age interval (ca. 3300 cal yr BC–middle 1st millennium AD) and such cultures as Zaisanovskaia, Yankovskaia, and Krounovskaia of the coastal zone of today’s northern North Korea and the RFE, which share several subsistence traits with the Okhotsk culture.
…
II. While establishing that the Okhotsk Culture and its people were responsible for cultivating barley, with broomcorn and fox millet as a complementary subsistence strategy to carrying on a hunter-fisher-gatherer lifestyle, Leipe et al. 2017 (see II above) ultimately pinpoint the origin of barley introduction as issuing from the continental Russian Far East. This picture of barley introduction and dispersal routes could likely throw light upon the migration path taken by the Okhotsk people.

Lister et al.’s 2018 paper Barley heads East: Genetic analyses reveal routes of spread through the diverse Eurasian landscapes , illustrates that the dispersal routes (see map above) for barley are divergent, with a northern route passing Sakhalin into Hokkaido, and another southern one from East Asia into Japan. Leipe et al., 2017 had already established that there were two distinct genotypes of barley, the boundaries for the southern type lay at the Satsumon boundary in SW Hokkaido. This macro-view of barley dispersals shows that an interdisciplinary approach tying perspectives from archaeological investigations into agriculture with genomic research into barley genotypes, is also useful in illuminating the migratory path taken by specific prehistoric genetic lineages of cultures that have ostensibly disappeared from modern populations, and which can help explain the genetic landscape of current populations.b
…
III. Who are the Okhotsk culture people(genetic investigation approach)?
This segment attempts to tease out the genetic characteristics of the Okhotsk people at their point of entry into Hokkaido (analysis of remains at the Moyoro site) and correlate them with other anthropological traits.
Introduction via Sato et al., Mitochondrial DNA haplogrouping of the Okhotsk people based on analysis of ancient DNA: an intermediate of gene flow from the continental Sakhalin people to the Ainu, 2009 Anthropological Science
The Okhotsk culture developed around the southern coastal regions of the Okhotsk Sea during the 5th–13th centuries (Amano, 2003a). The Okhotsk culture differs in certain respects from the Epi-Jomon culture (3rd century BC–7th century AD) and the Satsumon culture (8th–14th centuries: Amano, 2003b), which were contemporary with the Okhotsk culture and developed in the southern and inner parts of Hokkaido Island. A particular feature of the Okhotsk culture is adaptation of their lifestyle to sea fishing and hunting. Therefore, archaeological sites of the Okhotsk culture are restricted to coastal regions. Moreover, polygonal large houses and rituals using animals, including brown bears, which are other typical features of the Okhotsk culture, have not been observed in the Epi-Jomon and Satsumon cultures. In addition, skulls of the Okhotsk people share particular morphological characteristics, which are anthropologically clearly different from those of the Epi-Jomon and Satsumon people. Hence, there has been much discussion among archaeologists and anthropologists as to the origins of the Okhotsk people. Morphological studies have revealed that the characteristics of the Okhotsk people are similar to those of the Nivkhi and Ulchi people, currently distributed around Sakhalin and the lower regions of the Amur River (Ishida, 1988, 1996; Kozintsev, 1990, 1992; Komesu et al., 2008). However, the origins of the Okhotsk people have not yet been clarified.
The closer archaeological relationships between the Ainu and Okhotsk cultures have also been investigated. Utagawa (2002) reported the occurrence of bear-sending ceremonies based on evidence obtained from archaeological sites of the Okhotsk culture. In the Ainu culture (17th century to the present day), the people consider brown bears to be a mountain god and perform a bear-sending ceremony called “Iomante” using juvenile bears nursed in the villages. These facts suggest that the Okhotsk people merged with the Satsumon people (a direct ancestoral lineage of the Ainu people) on Hokkaido, resulting in the establishment of the Ainu people (Utagawa, 2002).
Sato et al. (2007) analyzed the mitochondrial DNA (mtDNA) hypervariable region (HVR) 1 from bone remains of the Okhotsk people excavated from archaeological sites, and reported that the Okhotsk people were closely related to populations that are currently distributed around Sakhalin and the lower regions of the Amur River. In addition, Sato et al. (2007) suggested gene flow from the Okhotsk people to the Ainu people. …
(Because skulls excavated from the Moyoro site (Figure 1) lacked archaeological information, their dates were determined by the radiocarbon method (Yoneda et al., 2004). The dates were consequently estimated to be from the 7th to the 13th centuries AD, with the marine reservoir effect (400 14C years; Yoneda et al., 2007), in agreement with the Okhotsk culture period.)
Source: Sato et al., Mitochondrial DNA haplogrouping of the Okhotsk people based on analysis of ancient DNA: an intermediate of gene flow from the continental Sakhalin people to the Ainu, 2009 Anthropological Science
The haplogroup frequencies in the Okhotsk specimens were as follows: A, 8.1%; B5, 2.7%; C3, 5.4%; G1, 24.3%; M7, 5.4%; N9, 10.8%; Y, 43.2% (Table 2). Thus, in the mitochondrial gene pool of the Okhotsk people, haplogroup Y was major. This genetic feature is similar to those of populations currently living around the lower regions of the Amur River, such as the Ulchi, Nivkhi, and Negidal (Table 2). Table 3 shows values of pairwise Fst, estimated by haplogroup frequencies, among northern Asian populations. The exact test demonstrated that differentiations in any pair of the populations were statistically significant (P< 0.05). The genetic relationships between the Okhotsk people and the other northeastern Asian populations were shown using the multidimensional scaling method (Figure 3). Based on this, the Okhotsk people were closely related to the Ulchi, Ainu, and Negidal. Although the Nivkhi also neighbored the Okhotsk people (Figure 3), the Fst values between the Nivkhi and Okhotsk people were 0.1556: this high value is probably due to a high frequency of haplogroup Y (66.1%) in the Nivkhi. The findings show that the Okhotsk people are genetically closer to populations currently living around the lower regions of the Amur River as well as to the Ainu people of Hokkaido.
Discussion
In the present study, 37 specimens of the Okhotsk people were assigned to mtDNA haplogroups. The most characteristic feature of the gene pool of the Okhotsk people is the high frequency of haplogroup Y (43.2%; 16 of 37 individuals, Table 1 and Table 2), followed by haplogroups G1b (24.3%), N9b (10.8%), and the others. Recent studies have shown that most people with haplogroup Y are currently distributed around Sakhalin, the lower regions of the Amur River, and in the Ainu on Hokkaido Island (Schurr et al., 1999; Kivisild et al., 2002). The Ulchi, Nivkhi, and Negidal shared haplogroup Y at relatively higher frequencies (37.9%, 66.1%, 21.2%, respectively, Table 2). In addition, Adachi et al. (2006) reported that haplogroup Y was not observed among the Jomon people of Hokkaido, which are archaeologically considered to be a direct ancestral lineage of the Ainu (Table 2).
The present study showed that the Okhotsk people are genetically closely related to the Ulchi, Negidal, and Ainu (Figure 3). In addition to these three populations, the Nivkhi shared haplogroup Y at high frequencies, suggesting a genetic affiliation with the Okhotsk people. Sato et al. (2007) reported that the Ainu were not clustered with the Okhotsk people in the neighbor-joining tree using net nucleotide diversities (da distances) between populations (Nei and Li, 1979), while the da distance between the Okhotsk people and the Ainu was smaller than those between the Okhotsk people and other populations except for the Nivkhi and Ulchi. These facts indicate that the Ainu are also closely related to the Okhotsk people, although the Ainu people have been considered to be one of descendants of the Jomon people on the Japanese islands. The results obtained in the present study suggest that the Okhotsk people genetically originated from people living around the lower regions of the Amur River and that gene flow from the Okhotsk people to the Ainu occurred (Figure 4), in agreement with the report of Sato et al. (2007). Tajima et al. (2004) reported occurrence of gene flow from the Nivkhi to the Ainu. In addition, Hanihara et al. (2008) reported some morphological association of the Ainu with continental populations of northeastern Asia. The present study and the previous study (Sato et al., 2007) strongly show that the Okhotsk people played a role as an intermediate in gene flow from the populations living in the lower regions of the Amur River to the Ainu.[with the dangers of partial sequences and small samples of earlier studies, this needs to be investigated further: could the gene flow/genetic exchanges have been bidirectional or multidirectional over different prehistoric times, obscuring or distorting the true source of peopling origins???]
On the other hand, some of the Okhotsk people shared haplogroup A, which is not seen in populations currently living around the lower regions of the Amur River and in the Jomon people of Hokkaido (Table 1, Table 2). Haplogroup A is shared by many northeastern Asian populations except for the Nivkhi, Ulchi, and Negidal. The HVR 1 sequence of haplogroup A observed among the Okhotsk people is shared by the Koryak living around the Kamchatka peninsula. This sequence of HVR 1 corresponds with Type 3 reported by Sato et al. (2007). In addition, the Koryak are closely related to the Okhotsk people (Figure 3). These suggest that gene flows between the ancient Koryak and Okhotsk people also occurred (Figure 4).

Haplogroups N9 and G1 were also major among the Okhotsk people (Table 1, Table 2). The two haplogroups are also major among northeastern Asian populations. Haplogroup N9 is shared by both of the Hokkaido Jomon people (65.9%) and the Udegey (30.4%) at high frequencies (Table 2). Meanwhile, haplogroup G1 is observed among native populations of the Kamchatka peninsula (68.1% in the Itelmen and 41.9% in the Koryak) at high frequencies (Table 2), and is also shared by the populations around the lower regions of the Amur River (10.3% in the Ulchi; 5.4% in the Nivkhi; and 27.2% in the Negidal) and the Hokkaido Jomon people (13.6%). These facts suggest that haplogroup frequencies of the Okhotsk people were increased as a result of interactions with neighboring northeastern Asian populations.
In conclusion, mtDNA haplogrouping in the present study demonstrated that the Okhotsk people were closely related to populations living around lower regions of the Amur River and the Ainu. This finding indicates that the Okhotsk people could have originated around[or could have migrated to the region from further north] the lower regions of the Amur River and must have merged with the ancestors (the Epi-Jomon and/or Satsumon people) of the Ainu. This supports the results of the HVR 1 analysis (Sato et al., 2007) and the morphological analyses (Ishida, 1988, 1996; Kozintsev, 1990, 1992; Komesu et al., 2008). Moreover, the present study indicates that the Okhotsk people were also affected by the gene flow from the Kamchatka peninsula. To further clarify the gene flow relating to the Okhotsk people, genetic information on the paternal and biparental gene lineages would be very useful.
H. KAZUTA ET AL. Genotype frequencies of the ABCC11 gene in 2000–3000-year-old human bones from the Epi-Jomon and Jomon sites in Hokkaido, Japan 2010, ANTHROPOLOGICAL SCIENCE
Figure 1. Geographical locations of archaeological sites where bone specimens analyzed in the present study were excavated. Circles, triangles and squares indicate archaeological sites of the Jomon, Epi-Jomon and Satsumon cultures, respectively.
Amano (2003b) reported that earthenware and pit houses at the end of the Okhotsk period seemed to have been strongly influenced by the Satsumon culture. In addition, ancient mtDNA analyses of the Okhotsk people (Sato et al., 2007, 2009a) revealed that haplogroup Y, which has not been identified from Jomon and Epi-Jomon specimens, was shared by the Okhotsk people at high frequency and by the Ainu at lower frequency. These findings suggest that the Okhotsk people merged with direct descendants of the Jomon people, resulting in the establishment of the Ainu….
For autosomal DNA of the Jomon/Epi-Jomon and Okhotsk people, Sato et al. (2009b) analyzed allele frequencies of the ABCC11 gene, and reported that the frequencies of recessive allele A increased in the order Jomon, Ainu and Okhotsk. In addition, they suggested that gene flow occurred from the Okhotsk people to the Jomon lineage; however, Sato et al. (2009b) succeeded in analyzing only one Epi-Jomon specimen, and information on the ABCC11 gene alleles of the Epi-Jomon people is still lacking. The ABCC11 gene determining earwax phenotypes is a particular gene providing information on the phenotypes of ancient people. The frequency of wet earwax in the Ainu is higher than among other northeastern Asian populations (Yoshiura et al., 2006). This high frequency of wet earwax in the in the wild type, and Δ27 indicates the 27 bp deletion.
The exact test showed that no deviation from Hardy– Weinberg equilibrium was observed in the Jomon people (P > 0.05). Concerning the Epi-Jomon people, the deviationfrom Hardy–Weinberg equilibrium was not observed for the SNP (P > 0.05), but was observed for the 27 bp deletion.The latter deviation could have been caused by the existence of one homozygote of the deletion in the Epi-Jomon specimens.
Allele frequencies of the ancient people of Hokkaido were compared with the previously reported data from modern East Asian populations (Table 2, Figure 2): 58 Ainu, 126 Nagasaki, 58 Okinawa, 30 Yonaguni, 52 Han Chinese, and 99 Korean (Yoshiura et al., 2006). The frequencies of recessive allele A in the Jomon and the Epi-Jomon people were 0.476 and 0.591, respectively. The frequencies of allele A increased in the order Jomon, Epi-Jomon, Ainu (0.759), and considered to have resulted from allele frequencies of the Jomon people in Hokkaido. Therefore, further analysis of the ABCC11 gene could provide new information for verification of the hypothesis on the migration and history of ancient people…
Results
The exact test showed that no deviation from Hardy– Weinberg equilibrium was observed in the Jomon people (P > 0.05). Concerning the Epi-Jomon people, the deviation from Hardy–Weinberg equilibrium was not observed for the SNP (P > 0.05), but was observed for the 27 bp deletion. The latter deviation could have been caused by the existence of one homozygote of the deletion in the Epi-Jomon speci- mens.
Allele frequencies of the ancient people of Hokkaido were compared with the previously reported data from modern East Asian populations (Table 2, Figure 2): 58 Ainu, 126 Nagasaki, 58 Okinawa, 30 Yonaguni, 52 Han Chinese, and 99 Korean (Yoshiura et al., 2006). The frequencies of recessive allele A in the Jomon and the Epi-Jomon people were 0.476 and 0.591, respectively. The frequencies of allele A increased in the order Jomon, Epi-Jomon, Ainu (0.759), and Okhotsk people (0.839). Exact tests of population differentiation showed that allele frequencies of the ABCC11 gene between the Jomon and Okhotsk people, between the Jomon people and Ainu, and between the Epi-Jomon and Okhotsk people were significantly different (P < 0.05) (Table 3). On the other hand, allele frequencies between the Jomon and Epi-Jomon people, between the Epi-Jomon people and Ainu, and between the Okhotsk people and Ainu were not significantly different (P > 0.05) (Table 3).
Evidence of the bear-sending ceremony, which has been maintained in the Ainu culture, was found at archaeological sites of the Okhotsk culture (Utagawa, 2002). The earthen- ware and pit houses at the end of the Okhotsk period seem to be strongly influenced by the Satsumon culture (Amano, 2003b). In addition, mtDNA analyses indicated that haplogroup Y, which has not been found in the Jomon and Epi- Jomon people in Hokkaido, was identified in the Okhotsk people at higher frequencies (Sato et al., 2007, 2009a). Haplogroup Y was also found from the Ainu at a relatively lower frequency (Horai et al., 1996). These three findings suggest that gene flow occurred from the Okhotsk people to the Satsumon people, who are considered to be direct ancestors of the Ainu. Because haplogroup Y has not been detected from the Epi-Jomon specimens (Adachi et al., 2006, 2009), it has been thought that gene flow might have occurred in the end of the Satsumon period.
The present study, however, show that the allele A frequency in the Epi-Jomon people (0.591) was higher than in the Jomon people (0.476), although the difference was not statistically significant. This difference in allele frequencies between the Jomon and Epi-Jomon people might have been caused by slight genetic influences of population(s), such as the Okhotsk people, who possessed a high frequency of al- lele A, or genetic drift in Hokkaido after the Jomon period.
A morphological study reported that the diversity of the Epi-Jomon people markedly increased from that of the Jomon people (Ishida, 1999); however, the exact test of population differentiation showed that ABCC11 allele fre- quencies between the Jomon and Epi-Jomon people were not significantly different (P > 0.05) (Table 3). Because this non-significant P value may have been caused by the small number of Epi-Jomon specimens successfully analyzed in the present study or the slight difference in allele frequencies between the Jomon and Epi-Jomon people, further investi- gation of the ABCC11 gene for additional Epi-Jomon speci- mens is necessary to clarify the allele frequency in the larger gene pool of the Epi-Jomon people. Masuda et al. (2001) re- ported that some juvenile (less than one year old) ancient brown bears (Ursus arctos) excavated from the archaeologi- cal site of the Okhotsk culture on Rebun Island of northern Hokkaido could have originated from southern Hokkaido occupied by the Epi-Jomon culture. In addition, Masuda et al. (2001) showed the intercultural association between the Okhotsk people of Rebun Island and the Epi-Jomon people of southern Hokkaido through the usage of juvenile bears for the bear-sending ceremony; therefore, genetic exchanges between the Epi-Jomon and Okhotsk people might have also occurred.
Otherwise, the results of the present study suggest that Yayoi immigrants to southern Hokkaido might have slightly affected genetically the Jomon people native to Hokkaido. In fact, Matsumura (2001) examined the dental characteristics of the ancient people of the Japanese archipelago, and reported that several Epi-Jomon specimens in southern Hokkaido were classified as immigrant types of the Yayoi or Okhotsk.
Perhaps going furthest with important implications for understanding the peopling dynamics and population substructure of Japan, and building upon previous genetic analyses, see: Adachi et al., who published the 2017 Ethnic derivation of the Ainu inferred from ancient mitochondrial DNA data where the authors examine the Edo Ainu and the known ancient DNA of Hokkaido Jomon, Epi-Jomon and Satsumon for their genetic components, comparing them with that of neighboring populations. Through their population cluster analysis, they have teased out hitherto unknown lineages of ancestry and clarified the relationships and genetic affinities of various populations, including the Okhotsk culture and other neighboring Siberian populations, and ultimately pinpointing the likely originating populations that have formed part of the ancestry or admixed with the earlier settlers of Hokkaido and southern islands of Japan.
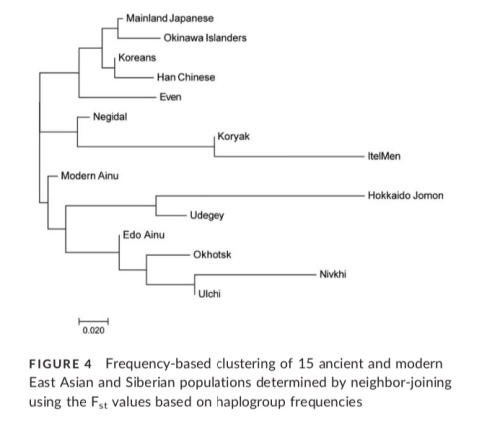
“the Ainu are considered to be descended from the Jomon people, with little admixture with other populations.
However, the results of our study suggest that the later admixture of Ainu with other populations than Jomon people was more considerable than it was proposed until today.
First, our results showed that the genetic influence of the Okhotsk culture people on the Ainu is significant. The proportion of Okhotsk-type haplogroups in the Edo Ainu was 35.1%, which is as high as that of the Jomon-type haplogroups (30.9%). This suggests that the Okhotsk culture people were one of the main genetic contributors to the formation of the Ainu.
Moreover, intriguingly, a genetic contribution of the mainland Japanese to the Edo Ainu is evident (28.1%), which is almost as considerable as those of the Jomon and the Okhotsk culture people. As referred to above, conventionally, the genetic influence of the mainland Japanese on the Ainu is considered to have been limited until the Meiji government started sending settlers to Hokkaido as a national policy in 1869. However, our findings cast doubt on this accepted notion.
In addition, Siberian-type haplogroups are observed in the Edo Ainu. Although their frequency is low (7.3%), as described earlier, the existence of these haplogroups may hint at the continuity of the genetic relationship between the Ainu and native Siberians …” Comparisons of the differing haplogroup components can be seen from their very helpful data below

See also Origins and genetic features of the Okhotsk people revealed by ancient mitochondrial DNA analysis Pdf fulltext found:
“…16 mtDNA haplotypes were identified from 37 individuals of the Okhotsk people. Of the 16 haplotypes found, 6 were unique to the Okhotsk people, whereas the other 10 were shared by northeastern Asian people that are currently distributed around Sakhalin and downstream of the Amur River. The phylogenetic relationships inferred from mtDNA sequences showed that the Okhotsk people were more closely related to the Nivkhi and Ulchi people among populations of northeastern Asia. In addition, the Okhotsk people had a relatively closer genetic affinity with the Ainu people of Hokkaido, and were likely intermediates of gene flow from the northeastern Asian people to the Ainu people. These findings support the hypothesis that the Okhotsk culture joined the Satsumon culture (direct descendants of the Jomon people) resulting in the Ainu culture, as suggested by previous archaeological and anthropological studies.”
“Molecular phylogenetic relationships between the Okhotsk people and modern Asian populations
To investigate the phylogenetic relationships between the Okhotsk people and modern Asian populations, the NJ relationships among the 17 Asian populations were con- structed (Fig. 3). In this phylogenetic tree, the Okhotsk people were clustered with the Nivkhi, Ulchi, Negidal, Koryak, and Even. Among them, the Nivkhi and Ulchi were much closer to the Okhotsk people, and clustered with more than 70% bootstrap values. The close relatedness among the three populations was in congruence with the high degree of sharing of mtDNA haplotypes. On the other hand, the Ainu people were phylogenetically distant from the Okhotsk people (Fig. 3). However, the dA distance
(0.068%, Table 3) between the Okhotsk people and the Ainu was smaller than those between the Okhotsk and other populations except the Nivkhi and Ulchi. Moreover, multidimensional scaling analysis (two-dimensional dis- play, Fig. 4) of the genetic relationships among the 17 Asian populations based on dA distances showed that the Nivkhi, Ulchi, Negidal and Ainu were much closer to the Okhotsk people than the other Asian populations. These findings demonstrate that the Okhotsk people are closely related to modern populations distributed around the Sakhalin and downstream of the Amur River as well as to the Ainu people of Hokkaido.
Discussion
Six haplotypes identified (types 2, 5, 6, 10, 15, 16) were specific to the Okhotsk people (Table 1). This finding shows that the Okhotsk people were genetically differen- tiated from other Asian populations. On the other hand, identification of the other ten haplotypes (types 1, 3, 4, 7, 8, 9, 11, 12, 13 and 14) from the Okhotsk people implies a close genetic relationship between the Okhotsk people and other northeastern Asian populations. In particular, the Okhotsk people had higher genetic affinities with the Nivkhi and Ulchi among the compared northeastern Asian populations (Figs. 3, 4). The Nivkhi and Ulchi populations are currently distributed in areas geographically close to the Okhotsk culture sites (Figs. 1, 2). Among haplotypes 12, 13 and 14 identified from 11 Okhotsk people, a unique combination of four transitional mutations (16189C- 16231C-16266T-16519C for types 12, 13 and 14) was shared and regarded as the motif sequence for mtDNA haplogroup ‘‘Y1’’ reported by Kivisild et al. (2002). Re- cent studies have shown that most people with haplogroup Y1 are distributed in northern Asia and Siberia (Schurr et al. 1999; Kivisild et al. 2002). Moreover, Adachi et al. (2006) reported that haplogroup Y occurred in the Ainu of Hokkaido but not in the Jomon people of Hokkaido. In the present study, we found that the Okhotsk people shared haplogroup Y1 at a similar frequency (30%, 11/37) to the Nivkhi (35%, 20/57) and the Ulchi (31%, 27/87) (Table 2). The result also suggests that the Okhotsk people are genetically closer to the Nivkhi and Ulchi because of the similarity in the frequencies of haplogroup Y1 between them. Moreover, haplogroup Y1 was shared also by the Ainu (6%, 3/51) (Table 2). That the frequency of Y1 is higher in the Okhotsk people (30%) than the Ainu (6%) suggests gene flow from the Okhotsk people to the Ainu. In addition, other haplotypes clarified to haplogroup Y1 were found: for example, two types from two individuals of the Ulchi, two types from six individuals of the Nivkhi, and one type from seven individuals of the Ainu. When these numbers are included, the same direction of gene flow is still apparent. Tajima et al. (2004) examined mtDNA phylogeny of modern Asian people (not including the Okhotsk people) and reported that there was gene flow from the Nivkhi to the Ainu. The present study demonstrates that the Okhotsk people were an intermediate in the gene flow from the Nivkhi to the Ainu.
In previous morphological studies, Mitsuhashi and Yamaguchi (1961) reported that skeletons of the Okhotsk people had morphological characteristics similar to those of northeastern Asian people. In addition, Yamaguchi(1974) and Ishida (1988, 1996) reported, based on cranial measurements, that the Okhotsk people were closer to populations that are currently distributed downstream of the Amur River. [Shared migrational edges confounding true source of genetic lineage origins]
[support from anthropological evidence] Moreover, Utagawa (2002) reported that evidence for the occurrence of bear-sending ceremonies, as seen in the Ainu culture, was found from archaeological sites of the Okhotsk culture. Rituals using brown bears are thought to be one proto-type of ‘‘Iomante,’’ which has been performed as a bear-sending ceremony in the Ainu culture. Therefore, archaeologists have generally assumed that the Okhotsk culture joined the Satsumon culture (eighth to fourteenth centuries; direct descendants of the Jomon people in Hokkaido) resulting in establishment of the Ainu culture. The direction of gene flow obtained in the present study is in agreement with the interpretation based on previous morphological and archaeological data. These facts demonstrate that the Okhotsk people could have originated from northeastern Asian populations such as the Nivkhi and Ulchi currently living around Sakhalin and downstream of the Amur River, and support the hypothesis that the Okhotsk people could have joined the Satsumon people resulting in the Ainu culture. [barley route suggests shared dissseminator further north]
Kikuchi (2004) reported that walrus tusks excavated from archaeological sites of the Okhotsk culture could have been brought from the ancient Koryak culture to the Okhotsk culture, because walrus are currently distributed in the Arctic sea and the Bering sea. In the present study, the Okhotsk people were found to have some genetic affinities with the Koryak and the Even living around the Kamchatka peninsula (Table 2; Fig. 3). These facts suggest that there were genetic and cultural exchanges between the Okhotsk people and the Koryak and Even.
In conclusion, the present study has provided new insights towards further understanding the phylogenetic landscape.”
…
IV. Viral markers have come of age, but are still vastly underutilized by anthropologists or geneticists to corroborate or fine-tune their conclusions.
see Disease and virus markers are useful in showing co-migration of peoples
where I review the past literature on the subject of JV Virus, BKV and HTLV migration markers and how they have shown population substructures which correlate with the findings of other studies in the field of Y- or MtDNA or autosomal DNA, as well as craniometric studies. They are, in many instances, more capable of pinpointing the origins of populations in specific geographical locations and regions.
V. Differing approaches creating greater ambiguity as well as clarity
What is particularly interesting to me is that perspectives derived from studies of other disease- such as virus-markers, HLA genes or blood markers, morphological cranial traits, individually may have inconclusive results but when interpreted together, will fill out the gaps of knowledge or add sufficient detail to corroborate the complex genetic landscape and population substructure seen from Adachi’s study.
Unique maternal genetic features of the Okhotsk people identified by ancient mtDNA analysis demonstrated their genetic differentiation and some genetic affinity with geographically neighboring modern populations such as the Nivkihi and Ulchi. The molecular phylogenetic data of the present study support the previous anthropological discussion based on morphological and archaeological data. For the next step, it will be necessary to investigate genetic features of paternal and biparental lineages using autosomal and Y-chromosomal markers for further clarification of the genetic origins of the Okhotsk people.
To illustrate, HLA gene studies had the effect of showing up shared haplotypes and typically find greater population substructure(Nakaoka et al., 2013) , revealing affinities between populations, but also introduced a lot of ambiguity in identifying source populations or even distinguishing admixture populations, when interpreting the results … as the following two studies on the Ainu show.
1. Jinam et al., 2012’s The history of human populations in the Japanese archipelago inferred from genome-wide SNP data with special reference to Ainu and Ryukyuan populations “showed in this study, however, the Ainu population seems to have experienced gene flow with two different populations, the Mainland Japanese and the yet unknown population in the north…”
“Now, all the CHB individuals are almost full of magenta ancestry component, and the Mainland Japanese and JPT individuals also consist of ∼30% magenta-colored ancestry component. There are two Japanese individuals, one in the Mainland Japanese and the other in HapMap JPT, who have >70% magenta component. These two individuals are also outliers in the PCA analysis shown in Figure 1a. If we consider blue- and magenta-colored ancestry components as the Jomon and the Yayoi factors, the intermediate orange-colored component is not easy to comprehend. It is possible that this does not correspond to a real ancestral population, but an artificially inferred component corresponding to the long-term admixture between the Jomon and the Yayoi genetic components. At k=4, the five outlier Ainu individuals observed in the PCA plot (those in red circles in Figures 1a and b) were differentiated from the rest of the Ainu, as indicated in the purple color. The Ryukyuan-specific ancestral component appeared at k=5. This is again not easy to interpret, and could be an artificially inferred component.
Generally speaking, the frappe results appear to be consistent with the PCA analysis in terms of the two patterns: (1) varying amounts of admixture in the Ainu with the Mainland Japanese and (2) the possible presence of another source population, which contributed to the genetic structure of the Ainu….
Possible mother population for the alternative admixture events with the Ainu population
Unlike admixture with the Mainland Japanese, it is difficult to ascertain the other potential source of admixture in the Ainu without a proper source population. Previous studies did support the idea of contact with Northern populations, which may have contributed to the genetic diversity in the Ainu. Archeological data point to an introduction of a distinct culture, which was quite different from the Satsumon culture, by the Okhotsk people into Hokkaido during the 7th–10th centuries.52The cultural contact with these northern peoples seemed to continue until recently. Genetic studies using mitochondrial DNA24 and human leukocyte antigen loci32 support this idea by showing close affinities between the Ainu and the Nivkhi who live in the Sakhalin Island and the Amur River region…”
2. See Bannai et al., 2000, Analysis of HLA genes and haplotypes in Ainu (from Hokkaido, northern Japan) supports the premise that they descent from Upper Paleolithic populations of East Asia. – NCBI:
A*0201, A*0206, A*2601, A*3101, B*1501, B*5101, B*3901, and B*3501 were observed at frequencies of more than 10% and most of these have previously been found in populations of not only Asians but also North and South American Indians. A*68012, which has not so far been detected in Hondo-Japanese, was found in the Ainu (3%). On the other hand, several alleles common in Hondo-Japanese, including HLA-A*3303, A*1101, B*4403, B*5201, B*5401, B*4601, and B*0702 were infrequent in Ainu (0-1%). Correspondence and neighbor-joining analyses of various populations based on HLA-A, -B and -DRB1 gene frequencies enabled distinction between Asian, Native South American, European, and African populations. The Ainu, as well as Tlingit (Na-Dene), were placed midway between other East Asians, including Hondo Japanese, and Native South Americans (Amerindians) in the correspondence analysis. Furthermore, several HLA-A-B and HLA-B-DR-DQ haplotypes common in the Ainu, are shared with some Native American populations. These observations strongly suggest a unique place for the Ainu as descendants of some Upper Paleolithic populations of East Asia, from whom some Native Americans may have descended.
My conclusion
Reconciling different types of data and the different perspectives/hypotheses from different disciplines in order to discern source populations and directionality of migrating lineages — is an agonizingly complex and mindboggling difficult task, given that the vastly different analytical approaches tend to arrive at opposing conclusions which are difficult to evaluate — as can be seen for example from — the contrarian views that the Ainu ancestors contributed to the surrounding Siberian populations but not the other way round, or — the elusive search for Austronesians in Japan. Notwithstanding those difficulties, evidence from interdisciplinary approaches must all the more be looked at, to make sense of otherwise ambivalent conclusions in research papers. The value of reconciling different types of data, evidence and perspectives is illustrated by the emergence of finer-resolution data, all serving to clarify potential migration routes and source populations, for example, the light thrown by Leipe, List and Adachi and others, upon the genetic contribution of the “extinct” prehistoric Okhotsk culture (for example, HLA gene or barley dispersal studies point to origins in Upp Paleolithic Siberians) and its genetic contribution to existing modern Ainu and Hokkaido populations.
